.
| Reptilia |
| †Parareptilia |
|
†Millerettidae 
|
|
| nimetön |
|
†Eunotosaurus
|
|
| †Hallucicrania |
|
†Lanthanosuchidae 
|
|
| †Procolophonia |
|
†Procolophonoidea 
|
†Pareiasauromorpha 
|
|
|
|
|
|
|
|
| Eureptilia |
|
†Captorhinidae 
|
|
| Romeriida |
|
†Paleothyris
|
†Paleothyris
|
.
|
|
| Diapsida |
|
†Araeoscelidia 
|
|
| Neodiapsida |
|
†Claudiosaurus
|
|
|
|
†Younginiformes 
|
|
| Reptilia |
| Lepidosauromorpha |
|
†Kuehneosauridae 
|
|
| Lepidosauria |
|
Rhynchocephalia (tuatarat. ja niiden sukupuuttoon kuolleet sukulaiset) 
|
Squamata (liskot ja käärmeet)  
|
|
|
|
| Archosauromorpha |
|
|
†Choristodera 
|
|
|
|
†Prolacertiformes 
|
|
|
|
|
†Trilophosaurus
|
|
|
|
.
|
†Rhynchosauria 
|
|
|
Archosauriformes (krokotiilit, linnut, dinosaurukset ja sukupuuttoon kuolleet sukulaiset)  
|
|
|
|
|
|
| Pantestudines |
|
†Eosauropterygia 
|
|
|
|
†Placodontia 
|
|
|
|
†Sinosaurosphargis
|
|
|
|
†Odontochelys
|
|
| Testudinata |
|
†Proganochelys
|
Kilpikonnat (kilpikonnat) 
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Systeemit
Kiertoelimistö

Lämpötilakuva monitoreista
Suurimmalla osalla matelijoistakin on kolme- ja-kolmikammioinen sydän, joka koostuu kahdesta eteisestä, yhdestä vaihtelevasti jakautuneesta kammiosta ja kahdesta aortasta, jotka johtavat systeemiseen verenkiertoon. Hapetetun ja hapettoman veren sekoittumisaste kolmikammioisessa sydämessä vaihtelee lajin ja fysiologisen tilan mukaan. Eri olosuhteissa hapeton veri voi ohjautua takaisin elimistöön tai hapellinen veri voi ohjautua takaisin keuhkoihin. Tämän verenkierron vaihtelun on oletettu mahdollistavan tehokkaamman lämmönsäätelyn ja pidemmät sukellusajat vesieläinlajeille, mutta sen ei ole osoitettu olevan fitness-etu.
Yleisestä fysiologiasta on joitakin mielenkiintoisia poikkeuksia. Esimerkiksi krokotiileilla on anatomisesti nelikammioinen sydän, mutta niillä on myös kaksi systeemistä aorttaa ja ne pystyvät siksi ohittamaan vain keuhkoverenkiertonsa. Myös joillakin käärme- ja liskolajeilla (esim. pytoneilla ja liskoilla) on kolmikammioinen sydän, joka muuttuu supistuksen aikana toiminnallisesti nelikammioiseksi. Tämän tekee mahdolliseksi lihasharju, joka jakaa kammion kammiodiastolen aikana ja jakaa sen kokonaan kammiosystolen aikana. Tämän harjanteen ansiosta jotkut näistä konnaeläimistä pystyvät tuottamaan kammioiden paine-eroja, jotka vastaavat nisäkkäiden ja lintujen sydämissä esiintyviä paine-eroja.
Hengitys
Hyönteisten keuhkot
Kaikki matelijat hengittävät keuhkojen avulla. Vesikilpikonnat ovat kehittäneet läpäisevämmän ihon, ja jotkin lajit ovat muokanneet kloaakkansa kasvattamaan kaasujenvaihtoon käytettävää pinta-alaa. Näistä sopeutumisista huolimatta hengitys ei koskaan täysin onnistu ilman keuhkoja. Keuhkotuuletus toteutuu eri tavoin jokaisessa matelijoiden pääryhmässä. Kyykäärmeillä keuhkot tuulettuvat lähes yksinomaan aksiaalisen lihaksiston avulla. Tämä on myös sama lihaksisto, jota käytetään liikkumisessa. Tämän rajoituksen vuoksi useimmat kyykäärmeet joutuvat pidättämään hengitystään intensiivisten juoksujen aikana. Jotkut ovat kuitenkin keksineet keinon kiertää se. Varanidit ja muutamat muut liskolajit käyttävät normaalia ”aksiaalista hengitystä” täydentämään buccal-pumppausta. Näin eläimet pystyvät täyttämään keuhkonsa täysin intensiivisen liikkumisen aikana ja pysyvät näin aerobisesti aktiivisina pitkään. Tegu-liskoilla tiedetään olevan protodiafragma, joka erottaa keuhkoontelon viskeraalisesta ontelosta. Vaikka se ei varsinaisesti kykene liikkumaan, se mahdollistaa keuhkojen suuremman täyttymisen, koska se vähentää sisäelinten painoa keuhkoista. Krokotiileilla on itse asiassa lihaksikas pallea, joka vastaa nisäkkäiden palleaa. Erona on se, että krokotiilien pallealihakset vetävät häpyluuta (lantion osa, joka on krokotiileilla liikkuva) taaksepäin, mikä laskee maksan alaspäin ja vapauttaa näin tilaa keuhkojen laajentumiselle. Tällaista pallea-asennetta on kutsuttu ”maksamännäksi”.
Kilpikonnat ja kilpikonnat

Punakorvakilpikonna hengittää
Kilpikonnien ja kilpikonnien hengittämistä on tutkittu paljon. Tähän mennessä vain muutamaa lajia on tutkittu niin perusteellisesti, että on saatu käsitys siitä, miten kilpikonnat sen tekevät. Tulokset osoittavat, että kilpikonnat ja kilpikonnat ovat löytäneet erilaisia ratkaisuja tähän ongelmaan. Vaikeutena on se, että useimpien kilpikonnien kuoret ovat jäykkiä eivätkä ne salli sellaista laajentumista ja supistumista, jota muut amniootit käyttävät keuhkojensa tuulettamiseen. Joillakin kilpikonnilla, kuten intialaisella läppäkilpikonnalla (Lissemys punctata), on keuhkoja ympäröivä lihaskalvo. Kun se supistuu, kilpikonna voi hengittää ulos. Lepotilassa kilpikonna voi vetää raajat sisään ruumiinonteloon ja pakottaa ilmaa ulos keuhkoista. Kun kilpikonna vetää raajojaan ulos, paine keuhkojen sisällä vähenee, ja kilpikonna voi imeä ilmaa sisäänsä. Kilpikonnien keuhkot ovat kiinnittyneet pintakuoren sisäpuolelle, ja keuhkojen alaosa on kiinnittynyt (sidekudoksen välityksellä) muihin sisäelimiin. Käyttämällä useita erityisiä lihaksia (jotka vastaavat suunnilleen palleaa) kilpikonnat pystyvät työntämään sisälmyksiään ylös- ja alaspäin, mikä johtaa tehokkaaseen hengitykseen, sillä monilla näistä lihaksista on kiinnityskohtia eturaajojen yhteydessä (monet lihakset laajenevat raajojen taskuihin supistuessaan). Hengitystä liikkumisen aikana on tutkittu kolmella lajilla, ja niiden hengitys on erilaista. Aikuiset naaraspuoliset vihreät merikilpikonnat eivät hengitä kainalossaan pesimärantoja pitkin. Ne pidättelevät hengitystään maalla liikkumisen aikana ja hengittävät jaksoittain levätessään. Pohjoisamerikkalaiset laatikkokilpikonnat hengittävät jatkuvasti liikkumisen aikana, eikä hengityssykli ole koordinoitu raajojen liikkeiden kanssa (Landberg ym., 2003). Ne käyttävät todennäköisesti vatsalihaksiaan hengittämiseen liikkumisen aikana. Viimeinen tutkittu laji on punakorvakilpikonna, joka myös hengittää liikkumisen aikana, mutta tekee pienempiä hengityshengityksiä liikkumisen aikana kuin pienten taukojen aikana liikkumiskertojen välillä, mikä viittaa siihen, että raajojen liikkeiden ja hengityslaitteen välillä voi olla mekaanisia häiriöitä. Laatikkokilpikonnien on myös havaittu hengittävän, kun ne ovat täysin sulkeutuneet kuorensa sisälle (ibid.).
Suupielet
Useimmilta matelijoilta puuttuu toissijainen suupieli, mikä tarkoittaa, että niiden on pidäteltävä hengitystä nielemisen aikana. Krokotiileilla on kehittynyt luinen toissijainen suulaki, jonka avulla ne voivat jatkaa hengittämistä pysyessään veden alla (ja suojata aivojaan kamppailevan saaliin aiheuttamilta vaurioilta). Myös skinkeillä (Scincidae-heimo) on vaihtelevassa määrin kehittynyt luinen toissijainen suulaki. Käärmeet valitsivat erilaisen lähestymistavan ja jatkoivat sen sijaan henkitorveaan. Niiden henkitorven jatke työntyy ulos kuin lihaisa olki, ja sen ansiosta nämä eläimet pystyvät nielemään suuria saaliita kärsimättä tukehtumisesta.
Nahka

Leguaanin takajalka, jossa näkyvät iguaanien ikoniset suomut.
Hyönteisten ihoa peittää kiimainen epidermis, joka tekee siitä vedenpitävän ja mahdollistaa matelijoiden elämisen kuivalla maalla, toisin kuin sammakkoeläinten. Nisäkkäiden ihoon verrattuna matelijoiden iho on melko ohut, eikä siinä ole paksua ihokerrosta, joka tuottaa nahkaa nisäkkäillä. Matelijoiden paljaat osat on suojattu suomuilla tai suomuilla, joilla on joskus luinen pohja ja jotka muodostavat panssarin. Liskojen ja käärmeiden kaltaisilla lepidosauruksilla koko iho on peittynyt päällekkäisten epidermaalisten suomujen peittoon. Tällaisia suomuja pidettiin aikoinaan tyypillisinä koko Reptilia-luokalle, mutta nyt tiedetään, että niitä esiintyy vain lepidosauruksilla. Kilpikonnien ja krokotiilien suomut ovat peräisin pikemminkin ihosta kuin epidermiksestä, ja niitä kutsutaan oikeammin suomuiksi. Kilpikonnien ruumis on piilossa kovassa kuoressa, joka koostuu sulautuneista suomuista.
Ekskreetio
Ekskreetio tapahtuu pääasiassa kahden pienen munuaisen avulla. Diapideilla virtsahappo on tärkein typpipitoinen jätetuote; kilpikonnat, kuten nisäkkäätkin, erittävät pääasiassa ureaa. Toisin kuin nisäkkäiden ja lintujen munuaiset, matelijoiden munuaiset eivät pysty tuottamaan nestemäistä virtsaa, joka on konsentroituneempaa kuin kehon neste. Tämä johtuu siitä, että niistä puuttuu Henlen silmukaksi kutsuttu erikoistunut rakenne, joka on olemassa lintujen ja nisäkkäiden nefronissa. Tämän vuoksi monet matelijat käyttävät paksusuolta apuna veden takaisinimeytymisessä. Jotkut pystyvät myös ottamaan virtsarakkoon varastoitunutta vettä. Joillakin matelijoilla ylimääräiset suolat erittyvät myös nenän ja kielen suolaisten rauhasten kautta.
Ruuansulatuselimistö

Vesikäärme Malpolon monspessulanus syö liskoa. Useimmat matelijat ovat lihansyöjiä, ja monet syövät ensisijaisesti muita matelijoita.
Useimmat matelijat ovat lihansyöjiä, ja niillä on melko yksinkertaiset ja verrattain lyhyet suolet, sillä liha on melko helppo hajottaa ja sulattaa. Ruoansulatus on hitaampaa kuin nisäkkäillä, mikä johtuu matelijoiden alhaisemmasta aineenvaihdunnasta ja kyvyttömyydestä jakaa ja pureskella ravintoa. Koska ne ovat poikilotermisiä (niiden ruumiinlämpö vaihtelee ympäristön mukaan), niiden energiantarve on noin viidenneksen tai kymmenesosan samankokoisen nisäkkään energiantarpeesta. Suuret matelijat, kuten krokotiilit ja suuret kuristimet, voivat elää yhdestä suuresta ateriasta kuukausia, koska ne sulattavat sen hitaasti.
Nykyaikaiset matelijat ovat pääasiassa lihansyöjiä, mutta matelijoiden varhaishistorian aikana useat ryhmät tuottivat kasvinsyöjämegafaunan: paleotsooisella kaudella pareiasaurukset ja synapsidiset dikynodontit ja mesotsooisella kaudella useat dinosaurusten linjat. Nykyään kilpikonnat ovat ainoa pääasiallisesti kasvinsyöjäkilpikonnaryhmä, mutta useat agam- ja iguaanilinjat ovat kehittyneet elämään kokonaan tai osittain kasveilla.
Kasvisyöjäkilpikonnilla on samat pureskeluongelmat kuin kasvinsyöjänisillä nisäkkäillä, mutta nisäkkäiden monimutkaisten hampaiden puuttuessa useat lajit nielevät ruuansulatuksen avuksi kiviä ja pikkukiviä (niin sanottuja gastroliitteja): Kivet huuhtoutuvat mahalaukussa ja auttavat jauhamaan kasviainesta. Fossiilisia gastroliitteja on löydetty sauropodeista. Myös merikilpikonnat, krokotiilit ja merileguaanit käyttävät gastroliitteja painolastina, joka auttaa niitä sukeltamaan.
Hermosto
Matelijoiden hermosto sisältää saman perusosan kuin sammakkoeläinten aivot, mutta matelijoiden aivot ja pikkuaivot ovat hieman suuremmat. Useimmat tyypilliset aistielimet ovat hyvin kehittyneet tiettyjä poikkeuksia lukuun ottamatta, joista merkittävin on käärmeen ulkokorvien puuttuminen (keski- ja sisäkorvat ovat olemassa). Kallohermoja on kaksitoista paria. Lyhyen sisäkorvansa vuoksi matelijat käyttävät sähköistä viritystä laajentaakseen kuultavien taajuuksien valikoimaa.
Matelijoita pidetään yleisesti vähemmän älykkäinä kuin nisäkkäitä ja lintuja. Niiden aivojen koko suhteessa niiden kehoon on paljon pienempi kuin nisäkkäiden, enkefalisaatiokertoimen ollessa noin kymmenesosa nisäkkäiden aivojen koosta. Krokotiileilla on suhteellisesti suuremmat aivot, ja niillä on melko monimutkainen sosiaalinen rakenne. Suurempien liskojen, kuten monitoreiden, tiedetään osoittavan monimutkaista käyttäytymistä, myös yhteistyötä. Komodon lohikäärmeen tiedetään harrastavan leikkiä.
Näkeminen
Useimmat matelijat ovat päiväeläimiä. Näkö on tyypillisesti sopeutunut päivänvalo-olosuhteisiin, ja niillä on värinäkö ja kehittyneempi visuaalinen syvyyshavainto kuin sammakkoeläimillä ja useimmilla nisäkkäillä. Joillakin lajeilla, kuten sokeilla käärmeillä, näkö on heikentynyt. Joillakin käärmeillä on ylimääräisiä näköelimiä (sanan väljimmässä merkityksessä) infrapunasäteilylle (lämmölle) herkkien kuoppien muodossa. Tällaiset lämpöherkät kuopat ovat erityisen hyvin kehittyneitä kuoppakyykäärmeillä, mutta niitä on myös booilla ja pytoneilla. Näiden kuoppien ansiosta käärmeet pystyvät aistimaan lintujen ja nisäkkäiden ruumiinlämmön, minkä ansiosta kuoppakäärmeet voivat metsästää jyrsijöitä pimeässä.
Sukupuolinen lisääntyminen

Useimmat matelijat lisääntyvät sukupuolisesti, kuten tämä Trachylepis maculilabris -skinkki

Matelijoilla on amnioottiset munat, joilla on kova tai nahkamainen kuori, ja ne vaativat sisäistä hedelmöitystä.
Useimmat matelijat lisääntyvät suvullisesti, vaikka jotkut kykenevät suvuttomaan lisääntymiseen. Kaikki lisääntymistoiminta tapahtuu cloacan kautta, joka on pyrstön tyvessä sijaitseva ainoa ulostulo/sisäänkäynti, josta myös jätteet poistuvat. Useimmilla matelijoilla on paritteluelimet, jotka ovat yleensä sisäänvedetyt tai ylösalaisin ja joita säilytetään ruumiin sisällä. Kilpikonnilla ja krokotiileilla uroksella on yksi keskimmäinen penis, kun taas kyykkäeläimillä, kuten käärmeillä ja liskoilla, on pari hemipenejä. Tuatareilla ei kuitenkaan ole paritteluelimiä, joten uros ja naaras vain painavat kloakkejaan yhteen, kun uros erittää siittiöitä.
Vähemmistö matelijoista munii lapsivesimunia, jotka on peitetty nahkaisella tai kalkkipitoisella kuorella. Amnion, chorion ja allantois ovat läsnä alkion elinaikana. Toukkavaiheita ei ole. Vivipariteetti ja ovovivipariteetti ovat kehittyneet vain kyykäärmeillä, ja monet lajit, kuten kaikki boat ja useimmat kyykäärmeet, käyttävät tätä lisääntymistapaa. Viviparisuuden aste vaihtelee: jotkut lajit yksinkertaisesti säilyttävät munat juuri ennen kuoriutumista, toiset tarjoavat äidin ravintoa keltuaista täydentämään, ja taas toisissa lajeissa ei ole lainkaan keltuaista, vaan kaikki ravintoaineet saadaan nisäkkäiden istukkaa muistuttavan rakenteen kautta.
Sukupuolista lisääntymistä on havaittu kyykkäeläimillä kuudessa liskoheimossa ja yhdessä käärmeessä. Joillakin konnaeläinlajeilla naaraspopulaatio kykenee tuottamaan yksisukupuolisen diploidisen kloonin emosta. Tätä suvuttoman lisääntymisen muotoa, jota kutsutaan parthenogeneesiksi, esiintyy useilla gekkalajeilla, ja se on erityisen yleistä teiideissä (erityisesti Aspidocelis) ja lacertideissä (Lacerta). Vankeudessa elävät komodolohikäärmeet (Varanidae) ovat lisääntyneet parthenogeneesin avulla.
Parthenogeneettisiä lajeja epäillään esiintyvän kameleonttien, agamidien, xantusiidien ja typhlopidien keskuudessa.
Joillakin matelijoilla esiintyy lämpötilariippuvaista sukupuolen määräytymistä (Temperature-dependent sex determination, TDSD), jossa haudontalämpötila määrää, kuoriutuuko tietystä kananmunasta koiras vai naaras. TDSD on yleisintä kilpikonnilla ja krokotiileilla, mutta sitä esiintyy myös liskoilla ja tuataroilla. Toistaiseksi ei ole varmistettu, esiintyykö TDSD:tä käärmeillä.
Puolustusmekanismit
Monet pienet matelijat, kuten käärmeet ja liskot, jotka elävät maassa tai vedessä, ovat alttiita kaikenlaisten lihansyöjäeläinten saalistukselle. Niinpä välttely on matelijoiden yleisin puolustautumismuoto. Ensimmäisessä vaaran merkissä useimmat käärmeet ja liskot ryömivät pois aluskasvillisuuteen, ja kilpikonnat ja krokotiilit syöksyvät veteen ja uppoavat pois näkyvistä.

Naamioitunut Phelsuma deubia palmun lehdellä
Hyönteiset voivat välttää yhteenottoja myös naamioitumalla. Käyttämällä erilaisia harmaita, vihreitä ja ruskeita sävyjä nämä eläimet voivat sulautua hämmästyttävän hyvin luonnollisen ympäristönsä taustaan.
Jos vaara syntyy niin äkillisesti, että pakeneminen voi olla haitallista, krokotiilit, kilpikonnat, jotkin liskot ja jotkin käärmeet sihisevät äänekkäästi kohdatessaan vihollisen. Kalkkarokäärmeet värisyttävät nopeasti hännän kärkeä, joka koostuu sarjasta sisäkkäisiä, onttoja helmiä.
Jos tämä kaikki ei pelota vihollista, eri lajit omaksuvat erilaisia puolustustaktiikoita.
Käärmeet käyttävät monimutkaista käyttäytymismallia hyökätessään. Jotkut nostavat ensin päänsä pystyyn ja levittävät kaulansa ihoa yrittäessään näyttää suuremmalta ja uhkaavammalta. Tämän epäonnistuminen voi johtaa muihin toimenpiteisiin, joita harjoittavat erityisesti kobrat, kyykäärmeet ja lähisukulaislajit, jotka käyttävät hyökkäykseen myrkkyä. Myrkky on modifioitua sylkeä, jota annetaan torahampaiden kautta.
Kun krokotiili on huolissaan turvallisuudestaan, se aukoo suunsa paljastaakseen hampaat ja keltaisen kielensä. Jos tämä ei onnistu, krokotiili kiihtyy ja alkaa tyypillisesti päästää sihiseviä ääniä. Tämän jälkeen krokotiili alkaa vakavoitua ja muuttaa asentoaan dramaattisesti saadakseen itsensä näyttämään pelottavammalta. Vartalo paisuu näennäisen koon kasvattamiseksi. Jos on aivan välttämätöntä, se voi päättää hyökätä vihollisen kimppuun.

Valkopäinen kääpiögecko, jolla on karistettu häntä
Jotkut lajit yrittävät purra, jotkin käyttävät päätään moukarina ja kirjaimellisesti murskaavat vastustajan, jotkin taas ryntäävät ryntäämään tai uivat uhkaajaa kohti kaukaa, jopa jahtaavat sitä maalle tai laukkaavat gallupilla perään.
Hännän vangiksi joutuvat gekot, skinkit ja muut liskot irrottavat osan hännän rakenteesta autotomiaksi kutsutun prosessin kautta ja pystyvät näin pakenemaan. Irronneen hännän heiluminen jatkuu, mikä luo harhaanjohtavan tunteen jatkuvasta kamppailusta ja vie saalistajan huomion pois pakenevasta saaliseläimestä. Eläin voi osittain uudistaa häntänsä viikkojen kuluessa. Uusi osa sisältää pikemminkin rustoa kuin luuta, ja iho voi olla selvästi värjäytynyt muuhun kehoon verrattuna.
Elävät matelijat
- Matelijat (vain elävät ryhmät)
- Luokka Crocodilia (krokotiilit, gaviilit, kaimanit ja alligaattorit): 23 lajia
- Luokka Sphenodontia (Tuatara Uudesta-Seelannista): 2 lajia
- Luokka Squamata (liskot, käärmeet ja amfisbaenidit (”matoliskot”): noin 7000 lajia.
- Luokka Testudines (kilpikonnat, kilpikonnat ja kilpikonnat): noin 300 lajia
-
-
-
Komodon lohikäärme, nukkumassa. Suurin elävä lisko
Kuvia lapsille
-
Liskot, Nouveau Larousse Illustré, 1897-1904: Huomaa sammakkoeläinten sisällyttäminen (krokotiilien alapuolella).
-
Maastrichtin kalkkikivilouhoksesta löydetty ”antidiluvialainen hirviö”, Mosasaurus, 1770 (aikalaiskaiverrus)
-
Ensimmäisillä matelijoilla oli anapsidityyppinen kallon katto, kuten nähdään permikautisessa Captorhinus-suvussa
-
Fylogeneettiset luokitukset ryhmittelevät perinteiset ”nisäkkäiden kaltaiset matelijat”, kuten tämän Varanodonin, muiden synapsidien joukkoon, ei elävien matelijoiden joukkoon.
-
Mesotsooinen kohtaus, jossa näkyy tyypillistä matelijoiden megafaunaa: dinosauruksia, mukaan lukien Europasaurus holgeri, iguanodontteja ja etualan puunrungolla istuskeleva Archaeopteryx lithographica.
-
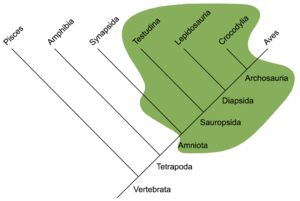
A = Anapsidi, B = Synapsidi, C = Diapsidi
-
Tyypillisen matelijan ja samankokoisen nisäkkään kestävä energiantuotto (jouleina) kehon sisälämpötilan funktiona. Nisäkkään huipputeho on paljon suurempi, mutta se pystyy toimimaan vain hyvin kapealla ruumiinlämpötila-alueella.
-
Hietaliskon iho, Näytetään squamate matelijoiden ikonisia suomuja
-
Kolubridikäärme, Dolichophis jugularis, syö jalkatonta liskoa, Pseudopus apodus. Useimmat matelijat ovat lihansyöjiä, ja monet syövät ensisijaisesti muita matelijoita.
-
Useimmat matelijat lisääntyvät sukupuolisesti, esimerkiksi tämä Trachylepis maculilabris -skinkki
-
Vuoden 1897 maalauksen taistelevasta ”Laelapsista” (nyk. Dryptosaurus) on tehnyt Charles R. Knight
-
Varhainen matelija Hylonomus
-
Varanus priscus oli jättiläismäinen petoeläimiä syövä vuohenpukin lisko, ehkä jopa 7 metriä pitkä ja jopa 1 940 kiloa painava.
|